- Information by Map Unit
- Information by Topic
- Documents in SoLo
Links to other related sites
- About SoLo
-
- Mission
- Disclaimer
- Authors (complete list)
The principal soil orders in western-montane forests are Inceptisols, Alfisols, Andisols, and Mollisols. Soil moisture and temperature regimes strongly influence forest type distribution and productivity. The most productive and resilient forests are on soils with udic moisture and frigid temperature regimes. Soils with low water holding capacity in ustic, xeric, and aridic moisture regimes and those with cryic temperature regimes are least productive and resilient. Soil organic carbon and nitrogen contents range from about 20,000 to more than 100,000 and 1,200 to 9,000 pounds per acre.
The western-montane forests occupy a vast area of the western United States in which active mountain building, erosion processes,and weathering regimes interact to produce landscapes and ecosystems of great diversity, grandeur, and degrees of resilience to land disturbance. Geologically, land surfaces vary from very recent (several thousands of years) to very old (millions of years). Soil-forming processes operating on these diverse landscapes have formed heterogeneous soils with highly variant properties. The resulting soil-forest ecosystems are likewise heterogeneous in character and function and require site-specific understanding for making management interpretations that provide for ecosystem sustainability and health. Nevertheless, some important principles and concepts can be established to serve as guides for forest resource management.
Soil is the primary medium for regulating movement and storage of energy and water and for regulating cycles and availability of plant nutrients. Soil also provides anchorage, aeration, heat for roots, and is home for many decomposers and element-transforming organisms. Informed inquiry and understanding are critical for making sound decisions about effective and efficient use and management of these vital resources. The objectives of this paper are to: (1) characterize the dominant soil-formation processes and properties in the principal western-montane forest types and landscapes; (2) illustrate the major soil moisture and temperature regime gradients of these forest types; and (3) discuss some implications for ecosystem function, productivity, and management.
The focus of this paper is on the western-montane forest types as described by Burns (1983) and the soils that support them. The principal forest types, their extent, and symbolic codes are in table 1. For further information about each of these types, refer to Agriculture Handbook 445. Additional information about the forest types particular to the Inland Northwest and their productivity, processes, and properties is provided by Harvey and others (1989).
The most direct way to characterize the kinds of soil occurring in western-montane forests is to discuss the occurrence of soil orders. Soil orders are the highest taxonomic class in the United States System of soil classification (Soil Survey Staff (1975). They are differentiated by the presence or absence of diagnostic horizons or features that reflect differences in the dominant soil-forming processes that occurred. The recent amendment to the National Soil Taxonomy Handbook established a new soil order Andisols (Soil Survey Staff 1989), making 11 soil orders. Hausenbuiller (1978) provides a more Survey Staff 1989),description of soil orders and the other taxonomic levels. Within the vast western-montane region, nine of the 11 soil orders occur, though several are rare. Only the highly weathered and leached Ultisols and Oxisols apparently are absent. Data are unavailable for precise estimates of the extent of soil orders. However, a review of selected soil survey reports and interpretation of data from "Soils of the Western United States" (Washington State University 1964) permits a reasonable estimate of relative occurrence (table 2). Probably the major difference between current estimates and the 1964 report is the relatively low occurrence of Spodosols. Many of the soils that were formerly thought to be Spodosols are now classified as Andisols, Alfisols, and Inceptisols.
Inceptisols are a diverse group of soils of intermediate development in humid and subhumid regions. They are mostly friable soils with moderate to high levels of organic carbon. Though many have high rock fragment content (more than 35 percent by volume) and have cryic temperature regimes, they usually are moderately deep and deep and are relatively fertile. Many are on unstable and active fluvial slopes in the Idaho Batholith.
Alfisols occur on a variety of landscapes, but most are stable or meta-stable surfaces. They support a wide range of forest types. Alfisols have relatively light-colored surface horizons and the dominant feature is the presence of an enriched subsurface clay layer. Most clays are the expanding 2:1 layer types. Ion exchange capacities are moderate to high and the soils have relatively high fertility. These soils dominate the Colorado plateau in the southern part of the western-montane forests (Meurisse and others 1975).
Andisols are forming mostly in volcanic ejecta such as ash, pumice, cinders, and lava. These soils have a characteristic layer of volcanic ash or pumice, 14 inches (35 cm) to several feet thick over buried soils. These soils, and intergrades to other orders, are among the most productive of western-montane forest soils (Meurisse 1985, 1987). Andisols occur mostly in the northwestern and northern parts of the western-montane region, on plateaus and gentle to moderately steep mountain slopes. Some are on steep and very steep slopes in northern Idaho and have a high erosion hazard.
Mollisols are characterized by thick, dark-colored surface horizons with at least 1 percent organic matter in the surface mineral soil layers. They occur mostly in the relatively dry western-montane forests and often have an abundance of grass and forbs in the understory. Many of the forests transitional to pinyon-juniper forests and grasslands occur on these soils. They occur mostly on stable mountain slopes and high plateaus.
The Aridisols occupy the driest parts of the Region, and the principal forest is the pinjon-juniper type. Entisols are forming in parent materials of recent origin, in cold climates, and on some unstable land surfaces where erosion prevents development of diagnostic features. Spodosols are strongly acid and have low levels of base cations. They have predominantly cryic temperature regimes and support Engelmann spruce (Picea engelmannii), subalpine fir (Abies lasiocarpa), and lodgepole pine (Pinus contorta) forest types.
Available soil moisture and soil temperature regimes are two overriding variables affecting the distribution, species mix, and productivity of western-montane forests. Soil moisture and temperature regimes are internal soil properties and are described in the Soil Taxonomy Handbook (Soil Survey Staff 1975) and by Hausenbuiller (1978). Generalized descriptions of these regimes are in table 3. The distribution of the principal western-montane forest types, by soil moisture and temperature regimes, is illustrated in figures 1 and 2. The size of the delineation is proportional to the distribution of the forest type according to these regimes and mean annual precipitation. It does not necessarily relate to aerial extent. For example, the ponderosa pine (Pinus ponderosa)/Rocky Mountain Douglas-fir (Pseudotsuga menziesii var glauca) type is the most extensive, except for the pinyon-juniper type (table 1). Yet, its soil moisture and temperature regime and mean annual precipitation range is relatively narrow. It is mostly on ustic and frigid regimes, but some are mesic and cryic and some are udic. The mixed conifer/western white pine (Pinus monticola)/western red cedar (Thuja plicata) type is on neither extremely cold nor very dry soils. It is generally the most productive of all the types in the region and is mostly on Andisols and Alfisols. At culmination of mean annual increment (CMAI), growth rates usually range from about 85 percent to about 150 ft 3/acre/yr (Burns 1983; Donaldson and others 1982; Donaldson and others in press).
| 1Measured at 50cm (20 in.) depth | |
| Soil moisture regimes | |
|---|---|
| Name | Description |
| Xeric | The soil moisture control section is dry for 45 or more consecutive days within the 4 months after the summer solstice. Typified in Mediterranean climates where winters are moist and cool and summers are warm and dry. |
| Ustic | Soil moisture control section is dry for 90 or more cumulative days, but is moist in some part for more than 180 cumulative days. Soil moisture is limited, but available when conditions are suited for plant growth. |
| Udic | Soil moisture control section is not dry for as long as 90 days (cumulative) and not dry for as long as 45 consecutive days in the 4 months following the summer solstice. Stored soil moisture plus rainfall ≥ ET amount. |
| Aridic | Soil moisture control section is dry more than half the time that the soil temperature is above 41°F. Never moist for 90 consecutive days when soil temperature is above 47°F. |
| Soil temperature regimes | |
| Name | Description |
| Mesic | The mean annual soil temperature is 8°C (47°F) or higher, but lower than 1°C (59°F). The difference between mean summer and winter temperature is more than 5°C (9°F). |
| Frigid | The mean annual soil temperature is lower than 8°C (47°F) and the difference between the mean winter and summer soil temperature is more than 5°C (9°F). |
| Cryic | Soils have a mean annual temperature higher than 0°C (32°F) but lower than 8°C (47°F) and a mean summer temperature lower than 8°C (47°F) with an O Horizon. |
Figure 1—Distribution of western-montane forest types in ustic and udic soil moisture regimes as a function of mean annual precipitation, and mesic, frigid, and cryic soil temperature regimes. [Text description of this figure]
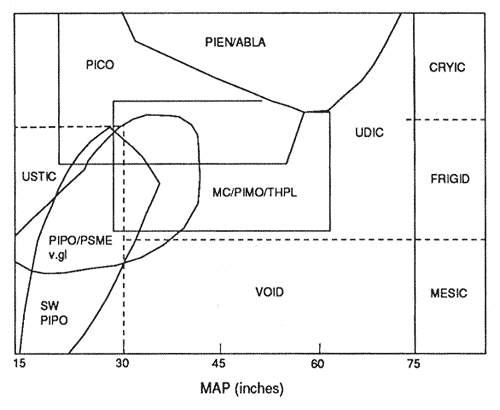
Figure 2—Distribution of western-montane forest types in xeric and udic soil moisture regimes, as a function of mean annual precipitation, and mesic, frigid, and cryic soil temperature regimes. [Text description of this figure]
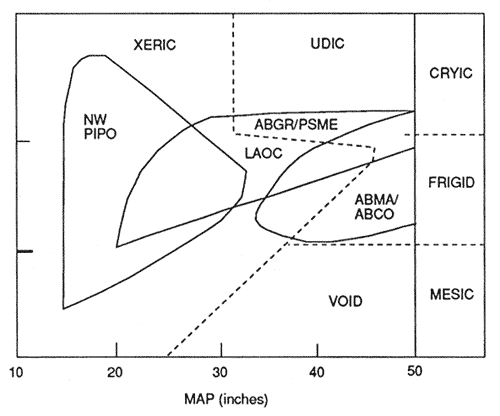
The grand fir (Abies grandis)/Douglas-fir and western larch (Larix occidentalis) types occur in xeric and udic moisture regimes and usually are on soils derived from volcanic ash (Andisols). Many of these soils, such as the Tolo series, currently are classified as having xeric moisture regimes. However, data from Klock (1980) and Geist and Strickler (1978), suggest that many of these soils probably have an udic moisture regime. When these soils are at elevations higher than 4,300 feet and mean annual precipitation is more than 40 inches, moisture regimes may be udic. Where there is little volcanic ash, less than 7 inches (18 cm), these forest types are largely on soils with xeric moisture regimes.
Temperature regimes are mostly frigid, but some are cryic. The Andisol and Andic intergrade soils in these forest types have high readily available water holding capacities (Geist and Strickler 1978). This is a major factor contributing to relatively high productivity of 65 to 125, or more, ft 3/acre/yr at CMAI (Meurisse 1985, 1987). The northwest ponderosa pine forest (NW PIPO) is clearly the dominant xeric regime type. Though not shown in figure 2, some of the lodgepole pine type in association with the NW PIPO type also is xeric. Much of the NW PIPO type is on soils with cryic temperature regimes. The cryic soils under the NW PIPO type are mostly the pumiceous Andisols of the central Oregon plateau (Carlson 1979; Meurisse 1985, 1987). These soils are known to exhibit unique thermal properties which affect their management (Cochran 1975). This type is also on some soils with mesic temperature regimes. Plant available water-holding capacity is an important factor governing productivity on soils with a xeric moisture regime.
The lodgepole pine and Engelmann spruce/subalpine fir types are dominantly on soils with cryic temperature regimes and with udic moisture regimes. Thus, soil temperatures are a major limiting variable for tree growth. The southwest ponderosa pine forest type is mostly on soils with frigid and mesic temperature regimes and ustic moisture regimes. Soil water often is limiting plant growth and plant available water-holding capacity of the soils is critical. Some of the southwestern ponderosa pine forest has udic moisture regimes and is the most productive in this type (Meurisse and others 1975).
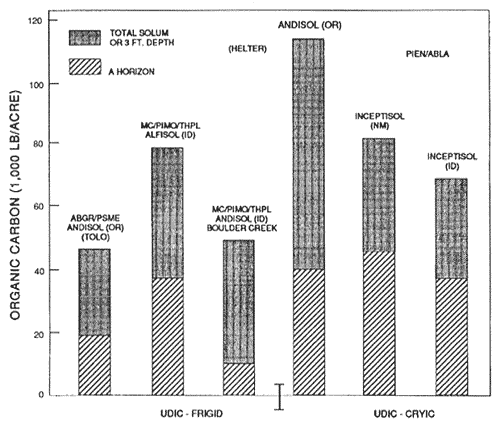
Soils form as the product of five major factors including climate, parent material, topography, organisms, and time (Jenny 1941). In western-montane forests, each of these factors assumes paramount importance in some part of the region. For example, continental and alpine glaciation have left mostly coarse-grained till deposits in the northern part of the region and at high elevations in the Rocky Mountains. Many soils from these parent materials have very dense layers, which are restrictive to both roots and water. The eruption of Mount Mazama, about 6,700 years before present, deposited volcanic ash, cinders, and pumice on much of the northern and western part of the region from the Cascades in Oregon and Washington to British Columbia, western Montana, and central to northern Idaho. Soils forming in this material, though very young, have very low bulk densities (0.5 to 1.0 g/cc) and have accrued relatively high amounts of organic matter (fig. 3). Thus, they are very favorable for root development and water movement and storage. The southern part of the region usually has very old surfaces of sedimentary origin. The resulting Alfisol are among the most weathered soils in the western-montane forests.
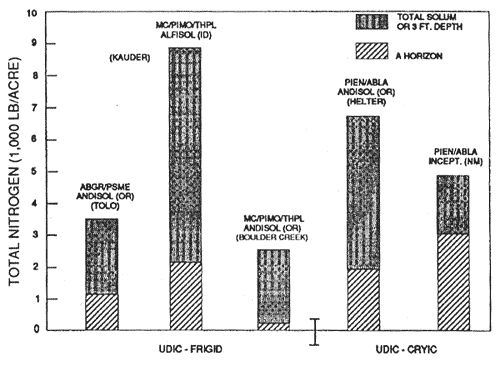
Figure 3—Mineral soil organic carbon content of representative soils and forest types in udic moisture and frigid and cryic temperature regimes. Source: Tolo and Helter adapted from Geist and Strickler, (1978); others from unpublished National Cooperative Soil Survey Laboratory data. [Text description of this figure]
The associated weathering processes of oxidation and reduction, hydration, mass flow, and ion exchange reactions result in soils with particular characteristics. However, soils are open-ended systems with many subprocesses and reactions. These processes of ongoing soil formation include: (1) additions to the soil; (2) losses from the soil; (3) transformations of material within the soil; and (4) translocations within the soil (Buol and others 1972). Table 4 illustrates many of these soil-forming processes which are active in western-montane forest soils. While many combinations, or even all, of these processes can occur to some degree in any soil, one or a few are dominant under specific conditions. Each soil order is the result of dominance by certain processes. For example, podzolization is dominant in Spodosols, but littering, illuviation, and eluviation also are important.
Losses from soils are from either erosion or leaching. In western-montane forests, leaching losses generally are minimal. Even in the temporary absence of forest cover, leaching losses seldom are important because of soils with relatively high ion exchange capacities and relatively dry soil conditions especially in the aridic, xeric, and ustic moisture regimes. Leaching losses may be important on some of the more moist sites with cryic temperature regimes, especially when vegetation is removed. Erosion losses are variable and can be accelerated with improper practices. This is particularly true on slopes greater than 40 percent and on soils with relatively low surface organic matter contents or moderate to low infiltration rates.
Additions to soils occur through littering, melanization, and cumulization. Wind-blown materials from adjacent arid regions represent a cumulization process that recycles bases such as calcium, magnesium, and potassium onto nearby forest soils. This gives rise to soils with moderate-to-high base status such as some Alfisols, Inceptisols, and Mollisols. In the absence of this process, some of these soils may advance in weathering stages to a degree that these nutrients would become limiting.
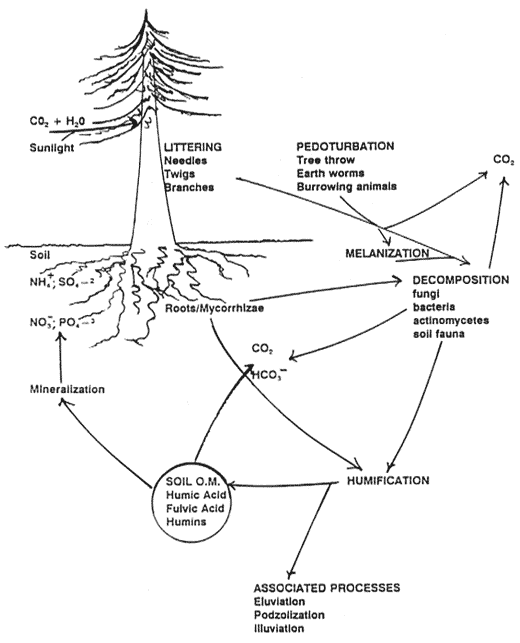
Littering, melanization, and pedoturbation are important processes in many soils, but especially in forested soils. Accumulation and mixing of organic matter are important for providing nutrients, ion exchange, aggregate stability, and water-holding capacity. Accumulation of organic matter also provides surface protection against raindrop splash, reducing the potential for erosion. Organic matter also serves as an energy source for organisms that perform many important functions in soils (Jurgensen and others 1979; Richards 1987). But these processes are only a part of the interaction of several processes that affect accumulation and cycling of carbon and nitrogen in soils. This includes the transformation process of mineralization, which is critical to making nitrogen and sulfur, and to a lesser extent phosphorus, available for plant growth (fig. 4). A thorough discussion of these processes in soil ecosystems, including the role of soil biota, is presented by Richards (1987). The principal source of soil organic carbon is from plant litter (detritus), roots, and decomposer organisms. Over half the soil organic matter comes from roots, mycorrhizae, and other fungi (Cromack and others 1979; Sollins and others 1980).
Figure 4—Dominant soil formation processes involving carbon cycling and accumulation in forest soils. The mineralization process releases nitrogen, sulfur, and phosphorus from soil organic matter in forms available for plant use. [Text description of this figure]
The rates and magnitudes of organic carbon and nitrogen cycling and accumulation vary widely among the many types of soils in western-montane forests. Some examples of carbon and nitrogen accumulation for representative soils are in figures 3 and 5 through 7. These processes are subject to both natural (fire, wind, volcanic eruptions) and anthropogenic influences. Such influences can have temporal changes that either increase or decrease the process rates and magnitudes. Thus, it is important to gain an understanding of these processes and how they function in given soil ecosystems in order to manage for sustainability of desired forest types.
Figure 5—Mineral soil organic carbon content of representative soils and forest types in xeric moisture and frigid and cryic temperature regimes. Source: Klicker adapted from Geist and Strickler (1960); Nevine from unpublished National Cooperative Soil Survey Laboratory data; LaPine from Dyrness (1960). [Text description of this figure]
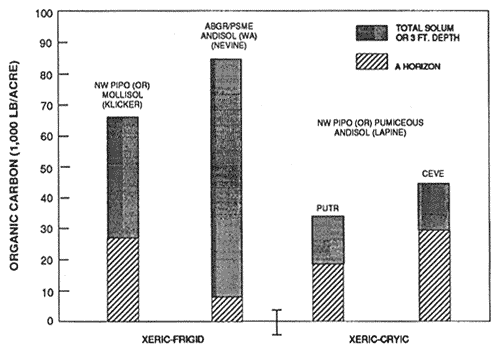
Figure 6—Mineral soil organic carbon content of representative soils and P-J SW PIPO and PIPO/PSME, forest types in ustic and udic moisture and mesic and frigid temperature regimes. Adapted from unpublished National Cooperative Soil Survey Laboratory data. [Text description of this figure]
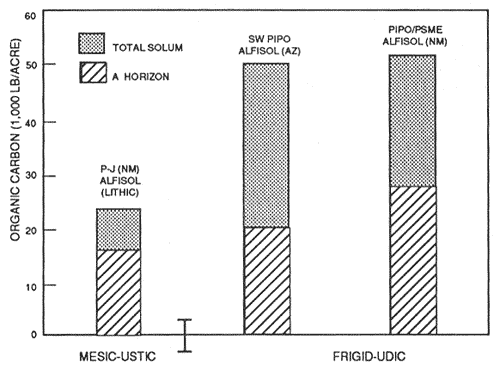
Figure 7—Mineral soil total nitrogen content of representative soils and forest types in udic moisture and frigid and cryic temperature regimes. Source: Tolo and Helter adapted from Geist and Strickler (1978); others from unpublished National Cooperative Soil Survey Laboratory data. [Text description of this figure]
In the most arid parts of the region, calcification is an important process. It occurs where soil moisture is insufficient to move calcium through the soil profile. Calcium accumulations are root restrictive, and tree roots seldom penetrate layers of calcium carbonate. While juniper appears to have low sensitivity, ponderosa pine is more sensitive. Thus, regeneration and tree growth are hampered.
To understand the function of soil properties in relation to productivity and management practices, it is necessary to understand concepts of productivity.
The Soil Science Society of America (1978) defines soil productivity as: "The capacity of a soil in its normal environment, for producing a specified plant, or sequence of plants, under a specified system of management."
Productivity is governed by the magnitude and rates of change of several state variables. The magnitudes and dynamics vary in space and time, are interactive among each other, and are affected by management practices both locally and globally. A general functional model of the state (fundamental) variables governing productivity: P=F (SM.SN.SA.L.H.V), where P is productivity as a measure of carbon fixation. Net primary productivity, though difficult to measure, is a complete measure of ecosystem dry-matter production. It is a useful measure for comparing different ecosystems (Grier and others 1989).
Other measures of practical use include site index and periodic increment per unit area. The state variables are: SM (available soil moisture), SN (available soil nutrients), SA (soil aeration), L (light quality and quantity), H (heat), and V (vegetation species and genetic traits). Site capacity factors (state variables) most easily affected by management activities are soil moisture, nutrients, and aeration. The inherent ecosystem properties of soil moisture and nutrient supply, aeration, and heat are a function of long-term soil processes and are principal productive function regulators. Many soil properties interact to affect each of these state variables. Productivity can increase, decrease, or remain unchanged depending on how the state variables are changed.
Several soil properties are paramount in their influence on available soil moisture, available soil nutrients, and aeration. These properties also are important in regional characterization of soils and in hydrologic processes.
Available Soil Moisture—Soil moisture regimes characterize the long-term general moisture supply. Properties such as soil depth, coarse fragment content, and organic matter content, and soil texture and porosity regulate water infiltration, movement, storage, and release of water to plants. For example, clay soils hold large quantities of water per unit volume, but they also hold water at very high energies so that availability to plants is restricted. At the other extreme, low-density, high-porosity volcanic ash and pumice soils hold high amounts of water that is readily available at low energies (Geist and Strickler 1979). Management practices that cause soil compaction and displacement reduce infiltration rates and soil volume. Thus, plant available water supply also is reduced. Shallow soils and skeletal soils (greater than 35 percent rock fragments) have lower available water supply than moderately deep and deep soils with few rock fragments.
In general, the soils with udic moisture regimes and Andisols have the highest potential to supply plant available moisture in western-montane forests. Plant available water holding capacity of these Andisols often is about 15 to 22 inches (Dyrness 1960; Geist and Strickler 1979). Table 5 illustrates some soil physical properties, including available water-holding capacity, for representative soil series or subgroups and their associated forest types. These soils are representative of Alfisols, Inceptisols, Andisols, and Mollisols and reflect the wide range of bulk density and available water-holding capacity of western-montane forest soils. Soils with inherent bulk density of 1.6 g/cc and greater usually restrict root development and penetration. Therefore, access to soil moisture is limited at those sites. Soils with these conditions usually are forming in dense glacial till or are some Alfisols that have fragipans or very high clay contents.
| Representative soil taxa | Forest type | Depth to restrictive layer (inches) | Surface bulk density (g/cc) | Subsoil bulk density (g/cc) | AWHC1 (inches) |
|---|---|---|---|---|---|
| 1Plant available water-holding capacity. | |||||
| Eutric Glosoboralf (AZ) | PIPO/PSME | ≥ 60 | 1.2 | 1.8 | 8–10 |
| Mollic Eutroboralf (AZ) | SW PIPO | 40–60 | 1.3 | 1.4 | 8–10 |
| Lithic Haplustalf (AZ) | P-J | < 20 | 1.2 | 1.3 | 0.9–1 |
| Dystric Cryochrept (NM) | PIEN/ABLA | 40–60 | 1.4 | 1.3 | 2–3 |
| Vay Series (ID) | PIEN/ABLA | 40–60 | .9 | 1.6 | 5–6 |
| Boulder Creek Series (ID) | MC/PIMO/THPC | ≥ 60 | .8 | .9 | 7–8 |
| Kauder Series (ID) | MC/PIMO/THPC | 34–40 | .9 | 1.7 | 8–10 |
| Tolo Series (OR) | ABGR/PSME | ≥ 60 | .7 | .8 | 15–22 |
| Nevine Series (WA) | ABGR/PSME | 40–60 | .6 | .8 | 6–7 |
| Nard Series (WA) | ABGR/PSME | ≥ 60 | 1.6 | 1.7 | 12–13 |
| LaPine Series (OR) | NW PIPO | ≥ 60 | .6 | .5 | 15–22 |
| Klicker Series (OR) | NW PIPO | 20–40 | .9 | .9 | 3–4 |
Available Soil Nutrients—Nutrient supply to plants is a function of several soil properties, including volume of root occupancy, mycorrhizae-root relationships, nutrient reservoirs, ion exchange properties, nutrient balance, and acidity. Soil texture, clay mineralogy, and organic matter content strongly influence ion exchange capacities, nutrient reservoir, mycorrhizal populations, and nutrient balance. Most western-montane forest soils are moderately acid to mildly alkaline (pH range of 5.6 to 7.8) and have base saturation of at least 35 percent. There are some soils, such as the Vay series, with base saturation of less than 35 percent. Such soils may be very strongly acid (pH 4.5 to 5.5). These are mostly cryic soils where leaching is prominent. Under these conditions, nutrients can be limiting plant growth. Under moderately alkaline conditions (pH 7.9 to 8.4), which may occur in soils with aridic and some ustic moisture regimes, nutrients also may be limiting.
Cation exchange capacities vary widely, but generally are adequate to retain an abundant supply of cations. Cation exchange capacities of Alfisols, Mollisols, and Aridisols usually are the highest of western-montane forest soils and range from 10 to more than 50 meq/100g. Some soils with low cation exchange capacities, such as some ashy and pumicious materials, have high internal porosity and are able to retain cations even after severe wildfire (Grier 1975). Soils with sandy loam and coarser textures, often from glacial till, usually have cation exchange capacities less than 10 meq/100g. When these soils occur in udic moisture regimes, nutrient losses can be significant, especially when vegetation is removed.
Nitrogen is the nutrient required in greatest quantity by trees and its abundance in the soil is a major factor in site quality. Nitrogen is known to be limiting in many western-montane forest soils (Cochran and Vander Ploeg 1988; Moore 1988). Total nitrogen, though very important, is only part of the story. More important, its availability when root activity is high is critical. Thus, the process of mineralization, which is influenced by soil moisture and temperature, and their effects on bacterial activity, determines whether nitrogen is available for plant uptake in amounts needed (fig. 3). This process also determines availability of sulfur and, to lesser extent, phosphorus. Thus, the soil organic matter content and nitrogen, sulfur, and phosphorus availability are inextricably linked. Figures 4 through 9 illustrate the contents of soil organic carbon and nitrogen in representative soils with different moisture and temperature regimes and forest types. The contents are for the mineral soil portion of the ecosystem. Organic carbon and nitrogen contents vary widely in western-montane forest soils. Also, the proportion in the surface layer varies measurably. These values are compared with the very high contents of about 250,000 to 725,000 lb/acre (280,000 to 812,000 kg/ha) of organic carbon and 15,000 to 38,000 lb/acre (16,800 to 42,560 kg/ha) of nitrogen in A and B horizons of some deep coastal Oregon Andisols under western hemlock (Meurisse 1972; 1976).
It is important to note that subtle differences may occur in soils under the same forest type, but with the presence of nitrogen-fixing understory plants. For example, figures 5 and 7 illustrate the Lapine series under ponderosa pine with an understory of bitterbrush (Purshia tridentata) and snowbrush (Ceanothus velutinus). Snowbrush fixes nitrogen in symbiosis with a bacterium in amounts of about 10 lb/acre/yr to 70 to 100 lb/acre/yr (Harvey and others 1989; Wollum and Davey 1975). Bitterbrush reportedly fixes less than 1 lb/acre/yr (Dalton and Zobel 1977; Haines and DeBell 1980). Nitrogen and organic carbon contents of surface soils are measurably increased in the presence of some nonleguminous N-fixing shrubs. Review of the data in figures 4 through 9 reveals some general patterns of organic carbon and nitrogen content. Generally, Andisols, especially those that are ashy and medial, have the highest contents of organic carbon and nitrogen in western-montane forest soils. The proportion in the surface to the total is highest in the weakly developed pumiceous Andisols. The soils in udic moisture regimes and frigid and cryic temperature regimes usually have the highest organic carbon and nitrogen content. The soils from more southerly latitudes generally have the least amount of organic carbon and nitrogen. Soils in ponderosa pine forests, except for Mollisols, usually are among the lowest in organic carbon and nitrogen content. Those soils with a relatively large proportion of carbon and nitrogen in the surface, relative to the total, generally are less resilient and more sensitive to soil disturbance, especially soil displacement.
Figure 8—Mineral soil total nitrogen content of representative soils and forest types in xeric moisture and frigid and cryic temperature regimes. Source: Klicker adapted from Geist and Strickler (1978); Nevine from unpublished National Coop. Soil Survey Laboratory data; LaPine from Dyrness (1960) and unpublished National Cooperative Soil Survey data. [Text description of this figure]
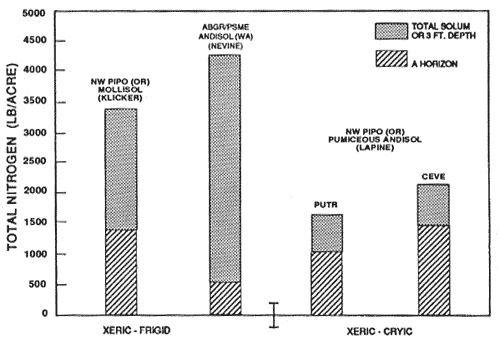
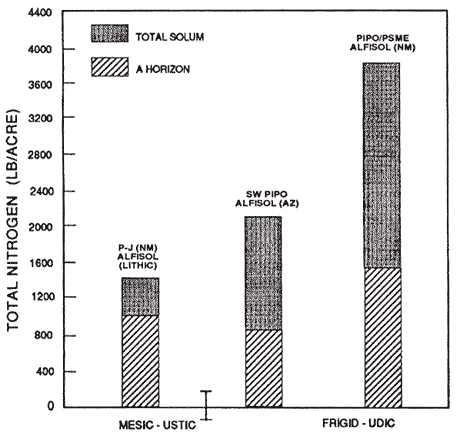
Figure 9—Mineral soil total nitrogen content of representative soils and P-J SW PIPO and PIPO/PSME forest types in ustic and udic moisture and mesic and frigid temperature regimes. Adapted from unpublished National Cooperative Soil Survey Laboratory data. [Text description of this figure]
Soil Aeration—The soil atmosphere or gaseous phase, is one of the three phases in nature represented in soil. The other two are soil matrix or solid phase and soil solution or liquid phase. Of the three, soil air is least discussed. Yet its importance in soil ecosystem function should not be underestimated. Soil aeration is needed for gas exchange with respiring roots. Aeration also effects soil biota and heat-transfer processes. Fortunately, most forest soils have high natural porosities, especially macro or air-filled pores and relatively low bulk densities (mass per unit volume). Indeed this is a characteristic of most temperate forests. However, soil porosity also is easily affected by forest management activities, especially where heavy ground equipment is used and concentrations of animals persist. Soil aeration is a function of soil porosity and drainage. The majority of western-montane forest soils are well drained or excessively drained and well aerated. Some soils with dense till layers or fragipans may have temporary or perched water tables with reduced aeration, but they are uncommon. Soil compaction and puddling from management practices is likely to have the greatest effect on aeration. The compaction process and effects in Pacific Northwest forests has been discussed by Froehlich and McNabb (1984). Similar effects can be expected in western-montane forests. Harvey and others (1989) discussed these processes and effects for mixed conifer and pine forests of the inland northwest where Andisols are common soils. Clayton and others (1987) found declines in one or more growth attributes in 15- to 25-year old ponderosa pine and lodgepole pine stands on Inceptisols with a volcanic ash surface layer. The declines were associated with increased bulk density, increased penetration resistance, and lateral soil displacement.
Soil bulk density is one of the most common indirect measures of porosity and soil strength in the field. Air permeameters have been used to measure macro porosity (Steinbrenner 1959). Soils with low bulk density have correspondingly high porosity. Data in table 5 illustrate some of the variation in soil bulk density for surface and subsoil horizons. While most of the soils have relatively low surface bulk density, several have very high subsoil density. Such soils, even though deep, have restricted root development. Soils with lowest densities are the Andisols and Andic intergrades. Some of the more dense and less porous soils are the Alfisols and other soils underlain by firm glacial till. The importance of low bulk density and high porosity on forest productivity and management of Andisols has been discussed by Meurisse (1985, 1987).
Soils are characterized by their composite set of properties and processes. The ability of soils to sustain long-term forest productivity is highly variable and is a function of their properties and processes. Similarly, soils vary in their resilience or ability to rebound or recover from management activities or natural disturbances. The diversity of soil types in western-montane forests suggests there is also diversity of resilience. The hazards of soil damage from erosion surface and mass failure compaction, puddling, and displacement often are governed by the shape and locations on the landscape, physical properties, and the kind and timing of activities. Indeed, some western-montane soils have very high hazards for erosion (Megahan and others 1978; Megahan 1981).
Nevertheless, many western-montane forest soils have properties that impart high resilience and low sensitivity (Meurisse 1985, 1987; Meurisse and others 1975). Some of the important properties affecting resilience are soil organic matter in both surface and subsoil layers, effective soil depth, inherent erodibility, plant available water-holding capacity, texture, structure, cation exchange capacity, shrink-swell potential, moisture regime, and temperature regime. To assess potential for soil damage, resilience, and productivity potential it is necessary to have high-quality soil surveys and to make site-specific soil investigations by qualified soil scientists. In doing so, the full potential of the many kinds of western-montane forest soils can be realized and their production capacity can be sustained for future generations. Also, for those soils with high sensitivity or low resilience, management prescriptions can be designed to overcome many of the inherent limitations and avoid detrimental soil damage, which would require costly rehabilitation or loss of productive potential of our precious soil resources.
Western-montane forest soils and landscapes are highly variable. The soils are forming from the interaction of many processes and materials. Each soil possesses a characteristic set of properties and processes, which must be understood to assure their management for long-term sustainability. Nine of the 11 soil orders are present. Inceptisols, Alfisols, and Andisols, respectively, are most abundant and occur in about 70 percent of the area. Soil-moisture regimes of western-montane forests are dominantly ustic, xeric, or aridic. Because these regimes are relatively droughty, available water-holding capacity of the soils is very important for forest productivity and reforestation. Several forest types are in udic moisture regimes where soil moisture generally is not the most limiting factor for plant growth. Rather, soil temperature in cryic regimes and soil fertility limit plant growth.
In the northern part of the region and at high elevations, continental and alpine glaciation have influenced soil formation. Where dense till layers occur, water movement and root growth are restricted. The eruption of Mount Mazama and subsequent deposits of volcanic ash, cinders, and pumice generally has enhanced the rooting medium. Protection of the surface layer of volcanic ash is critical for sustaining productivity of many of the soils in these forests.
Soil organic carbon and nitrogen contents vary widely among forest types and climatic regimes. Generally, the higher the organic carbon and nitrogen contents the higher potential productivity and soil resiliency.
Alexander, E. B. 1988. Morphology, fertility and classification of productive soils on serpentinized peridotite in California (USA). Geoderma. 41: 337-351.
Buol, S. W.; Hole, F. B.; McCraken, R. J. 1972. Soil genesis and classification. Ames, IA: Iowa State University Press. 354 p.
Burns, R. 1983. Silviculture systems for the major forest types of the United States. Agric. Handb. 445. Washington DC: U.S. Department of Agriculture, Forest Service. 191 p.
Carlson, G. 1979. Soil resource inventory, Winema National Forest. U.S. Department of Agriculture, Forest Service, Pacific Northwest Region, Winema National Forest. Klamath Falls, OR. 156 p. plus maps.
Clayton, J. L.; Kellogg, G.; Forrester, N. 1987. Soil disturbance-tree growth relations in central Idaho clearcuts. Res. Note. INT-372. Ogden, UT: USDA Forest Service, Intermountain Research Station. 6 p.
Cochran, P. H. 1975. Soil temperature and natural forest regeneration in south-central Oregon. In: Bernier, B; Winget C. H., eds. Forest soils and forest land management: Proceedings of the 4th North American Forest Soils Conference; 1973 August. Quebec, Canada: Laval University: 37-52
Cochran, P.; Vander Ploeg, J. 1988. Response results for other species fertilization trials (grand fir, ponderosa pine, western larch). Unpublished paper on file at: University of Idaho, College of Forestry, Wildlife and and Range Sciences; Intermountain Forest Tree Nutrition Cooperative, Moscow, ID.
Cromack, K.; Delwiche, C. C.; McNabb, D. H. 1979. Prospects and problems of nitrogen management using symbiotic nitrogen fixers. In: Gordon, J. C.; Wheeler, C. T.; Perry D. A., eds. Symbiotic nitrogen fixation in the management of temperate forests. Corvallis, OR: Forest Research Laboratory, Oregon State University: 210-223.
Dalton, D. A.; Zobel, D. G. 1977. Ecological aspects of nitrogen fixation by Purshia tridentata. Plant and Soil. 48: 57-80.
Donaldson, N.; De Francisco, J. T.; Barron, D. W. 1982. Soil survey of Stevens County, Washington, Spokane, WA: U.S. Department of Agriculture, Soil Conservation Service. In cooperation with U. S. Department of Agriculture, Forest Service, Washington Department of Natural Resources; Washington State University, Agricultural Research Center; and USDI Bureau of Indian Affairs. 459 p. plus maps.
Donaldson, N.; De Francisco, J. T.; Haagen, M. E.; Barron, D. W.; Coleman, R. G. [In press]. Soil survey of Pend Oreille County area, Washington. Spokane, WA. U.S. Department of Agriculture, Soil Conservation Service. In cooperation with U. S. Department of Agriculture, Forest Service; Washington Department of Natural Resources; and Washington State University, Agriculture Research Center.
Dyrness, C. T. 1960. Soil-vegetation relationships within the ponderosa pine type in the central Oregon pumice region. Corvallis, OR: Oregon State University. 217 p. Thesis.
Froehlich, H. A.; McNabb, D. H. 1984. Minimizing soil compaction in Pacific Northwest forests. In: Stone, E. L., ed. Forest soils and treatment impacts. Proceedings of the 6th North American Forest Soils Conference; 1983 June. Knoxville, TN: University of Tennessee: 159-192.
Geist, J. M.; Strickler, G. S. 1978. Physical and chemical properties of some Blue Mountain soils in northeast Oregon. Res. Pap. PNW-236. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 19 p.
Grier, C. C. 1975. Wildfire effects on nutrient distribution and leaching in a coniferous ecosystem. Canadian Journal of Forest Research. 5(4): 599-607
Grier, C. C.; Lee, K. M.; Nadkarni, N. M.; Klock, G. O.; Edgerton, P. J. 1989. Productivity of forests of the United States and its relation to soil and site factors and management practices: a review. Gen. Tech. Rep. PNW-222. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 51 p.
Haines, S. G.; DeBell, D. S. 1980. Use of nitrogen-fixing plants to improve and maintain productivity of forest soils. In: Impact of intensive harvesting on forest nutrient cycling: Proceedings of a symposium; 1979 August 13-16. Syracuse, NY: Syracuse University College of Environmental Science and Forestry: 279-303.
Harvey, A. E.; Larsen, M. J.; Jurgensen, M. F. 1981. Rate of woody residue incorporation into Northern Rocky Mountain forest soils. Res. Pap. INT-282. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 5 p.
Harvey, A. E.; Meurisse, R. T.; Geist, J. M.; [and others]. 1989. Managing productivity processes in the inland northwest-mixed conifer and pines. In: Perry, D. A.; Meurisse, R. T.; Thomas, B.; [and others], eds. Maintaining the long-term productivity of Pacific Northwest forest ecosystems. Proceedings of a symposium; 1987 spring. Corvallis, OR. Portland, OR: Timber Press. 256 p.
Hausenbuiller, R. L. 1978. Soil science, principles and practices. 2nd ed. Dubuque, IA: Wm. C. Brown Company. 611 p.
Jenny, H. 1941. Factors of soil formation. New York: McGraw-Hill. 281 p.
Jurgensen, M. F.; Larsen, M. J.; Harvey, A. E. 1979. Forest soil biology-timber harvesting relationships. Gen. Tech. Rep. INT-69. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 12 p.
Klock, G. O.; Lopushinsky, W. 1980. Soil water trends after clearcutting in the Blue Mountains of Oregon. Res. Note. PNW-361. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 8 p.
Megahan, W. F.; Day, N. F.; Bliss, T. M. 1978. Landslide occurrences in the western and central northern Rocky Mountain physiographic province in Idaho. In: Youngberg, C. T. ed. Forest soils and land use: Proceedings of the 5th North American Forest Soils Conference; 1978 August. Fort Collins, CO: Colorado State University: 116-139.
Megahan, W. F. 1981. Effects of silvicultural practices on erosion and sedimentation in the interior west-a case for sediment budgeting. In: Baumgartner, D. M., ed. Interior west watershed management: Proceedings of a symposium; 1980 April; Spokane, WA. Pullman, WA: Washington State University Cooperative Extension: 69-181.
Meurisse, R. T. 1972. Site quality of western hemlock and chemical characteristics of some western Oregon Andic soils. Corvallis, OR: Oregon State University. 164 p. Thesis.
Meurisse, R. T. 1976. Soil chemical and other properties of western hemlock soils in Oregon-their relationship to productivity. In: Atkinson, W. A.; Zazoski, R. T., eds. Western hemlock management: Proceedings of a conference, 1976 May; Alderbroh, WA. Contrib. No. 34. Seattle, WA: University of Washington, College of Forest Resources, Institute of Forest Products. 317 p.
Meurisse, R. T. 1985. Properties of Andisols important to forestry. In: Beinroth, F. H.; Lusio, W. L.; Maldonado, F. P.; Eswaran, H., eds. Proceedings of the Sixth International Soil Classification Workshop, Chile and Ecuador. Part 1: Papers. Washington, DC: U.S. Department of Agriculture, Soil Management Support Service: 53-67.
Meurisse, R. T. 1987. Forest soil management of the U.S. Andisols. In: Kimble, J. M.; Nettleton, D. N. eds. Proceedings of the First International Soil Correlation Meeting (ISCOM): Characterization, classification, and utilization of Andisols. Washington, DC: U.S. Department of Agriculture, Soil Management Support Services: 21-42.
Meurisse, R. T.; Leven, A. A.; Williams, J. A. 1975. Use potentials of the forested soils on the Colorado Plateau, Arizona. In: Bernier, B.; Winget, C. H., eds. Forest soils and forest land management: Proceedings of the 4th North American Forest Soils conference; 1973 August. Quebec, Canada: Laval University: 541-557.
Moore, J. 1988. Response results from the Douglas-fir nitrogen fertilization trials. Unpublished paper on file at: University of Idaho, College of Forestry, Wildlife and Range Sciences; Intermountain Forest Tree Nutrition Cooperative, Moscow, ID.
Richards, B. N. 1987. The microbiology of terrestrial ecosystems. New York: John Wiley and Sons. 399 p.
Soil Science Society of America. 1978. Glossary of soil science terms. Madison, WI: Soil Science Society of America. 36 p.
Soil Survey Staff. 1975. Soil taxonomy. a basic system of soil classification for making and interpreting soil surveys. Agric. Handb. 436. Washington, DC: U.S. Department of Agriculture, Soil Conservation Service. 754 p.
Soil Survey Staff. 1989. Soil Taxonomy. Amendments, Issue 13. Agric. Handb.436. Washington, DC: U.S. Department of Agriculture, Soil Conservation Service.
Sollins, P.; Grier, C. C.; McCorison, F. M.; [and others]. 1980. The internal element cycles of an old-growth Douglas-fir stand in western Oregon. Ecological Monographs. 50: 261-285.
Steinbrenner, E. C. 1959. A portable air permeameter for forest soils. Soil Science Society of America Proceedings. 23(6): 478-481.
Washington State University. 1964. Soils of the western United States. A regional publication by the Western Land Grant Universities and Colleges with cooperative assistance by the USDA Soil Conservation Service. Pullman, WA: Washington State University. 69 p.
Wollum, A. G.; Davey, C. B. 1975. Nitrogen accumulation, transformation, and transport in forest soils. In: Bernier, B.; Winget, C. H., eds. Forest soils and forest land management. Proceedings of the 4th North American Forest Soils Conference; 1973 August. Quebec, Canada: Laval University: 67-106.
Paper presented at the Symposium on Management and Productivity of Western-Montane Forest Soils, Boise, ID, April 10-12, 1990.
Robert T. Meurisse is Regional Soil Scientist, Pacific Northwest Region, USDA Forest Service, Portland, OR 97208. Wayne A. Robbie is Supervisory Soil Scientist, USDA Forest Service, Southwestern Region, USDA Forest Service, Albuquerque, NM. Jerry Niehoff and Gary Ford are Soil Scientists, Idaho Panhandle National Forests, USDA Forest Service, Coeur D'Alene, ID.